Prof. Efrat Monsonego Ornan
Ultra-processed food targets bone quality via endochondral ossification
https://www.nature.com/articles/s41413-020-00127-9
Ultra-processed foods are those food products which undergo repeated stages of processing and contain non-dietary ingredients. Culturally these foods are generally known as “junk food” and are typically pre-packaged and sold for mass consumption. The increasing prevalence of these products around the world have directly contributed to increased obesity as well as other mental and metabolic impacts on people across the globe- and of all ages. Much of the world has experienced a trend towards increased consumption of these products known to be easily accessible, relatively inexpensive and ready to eat out of the package. Children are known to be particularly large consumers of these products with estimates that as much as 70 percent of their caloric consumption comes from ultra-processed foods. And while many studies have reflected on the overall negative impact of these products, limited research has focused on their direct developmental effects on children- particularly in their younger years.
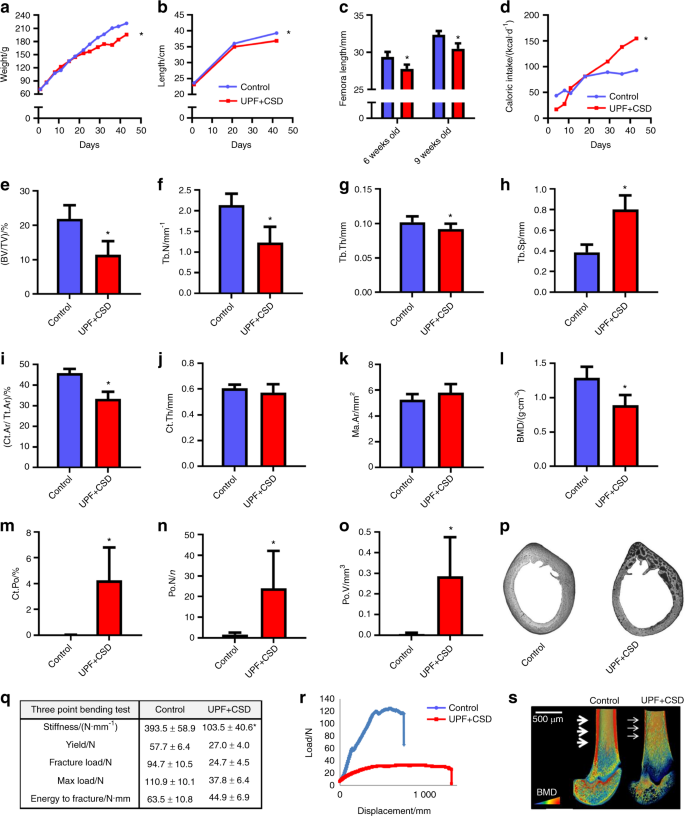
Our paper provides the first comprehensive analysis for how these foods impact skeletal development. The study surveyed lab rodents whose skeletons were in the post embryonic stages of growth. The rodents who were subjected to ultra-processed foods were shown to suffer from growth retardation and a direct impact on their bone strength in ways that were atypical for rodents of that age who would typically be healthy. Under histological examination, the researchers detected high levels of cartilage buildup in the rodents’ growth plates, the “engine” of bone growth.
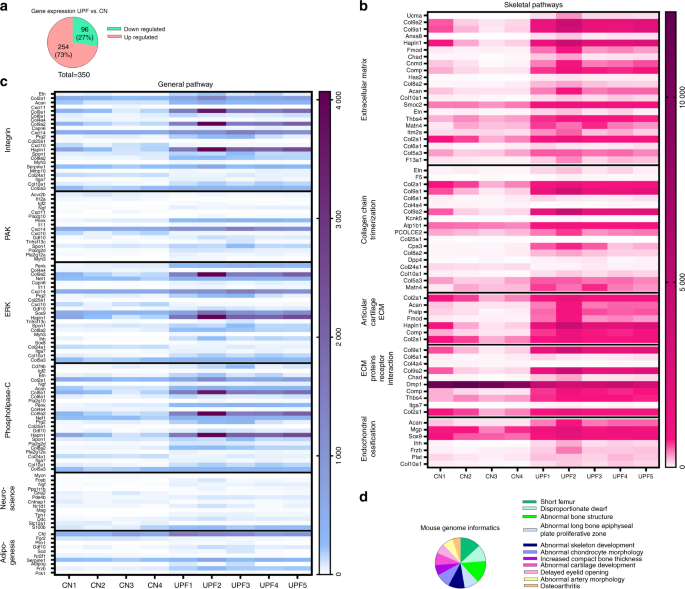
RNA sequence analysis of the growth plates demonstrated an imbalance in extracellular matrix formation and degradation and impairment of proliferation, differentiation and mineralization processes.
The next stage sought to investigate what specific ingredients in ultra-processed foods are suspected for impeding proper growth with the objective of determining whether increased fat and carbohydrate content are the damaging factors or perhaps decreased levels of calcium, phosphorus. While these decreased levels likely have negative impact on overall nutrition, they are not the main factors for explaining the highly damaging effect of these foods on the developing skeleton.

We then sought to analyze how specific eating habits might impact on people who consume these foods. Since 70 percent of a child’s food consumption comes from these foods, we worked to replicate this type of food intake. We divided the rodents weekly nutritional intake where thirty percent was from a ‘controlled’ diet and the remaining 70 percent consisted of ultra-processed foods. We found that the rodents experienced moderate damage to their bone density but lesser indications of cartilage buildup in the growth plates. Our conclusion was that even with a more limited intake, ultra-processed foods have a definite negative impact on skeletal growth.
The study serves as the first evidence of Ultra processed food direct impact on rodent skeleton development during the key stages of early growth and are believed to be even more damaging that the previous metabolic damage which is known to be associated with these products. These findings are critical because we know that children and adolescents consume these foods on a very regular basis to the extent that 50 percent of children in the US are ingesting them each and every day. Professor Carlos Monteiro, one of the world’s leading experts on nutrition has said that there is no such thing as a healthy ultra-processed food and he’s clearly right. Even if we reduce fats, carbs nitrates and other known harmful substances, these foods still possess their damaging attributes. Every part of the body is prone to this damage and certainly those systems that remain in the critical stages of development.
MECHANISMS GOVERNING SKELETAL DEVELOPMENT THROUGH ENDOCHONDRAL OSSIFICATION
The reason that vertebrates do not look like, say, jellyfish is their skeletal system, which supports their body and protects vital organs while enabling efficient mobility. These functions depend on a precise and optimally-designed overall body proportioning determined by the size of the skeleton, which in turn is determined by the rate and duration of bone growth. Longitudinal bone growth occurs via the chondrocytes at the growth plates (GP), through a very tightly regulated process of endochondral ossification. The chondrocytes differentiate and produce cartilage anlagen (endochondral) that is sequentially calcified, degraded and replaced by bone (ossification). When the system is functioning correctly, our legs are of equal length, as are our arms. However, disrupted regulation is manifested in skeletal diseases, malformations and height disarrays.
My interest in GP metabolism began while working towards my PhD, when my research dealt with the effect of growth hormone (GH) on chondrocyte proliferation and differentiation. After completing my PhD, I continued my training at the Weizmann Institute of Science by joining the effort to understand the basis for inherited human skeletal disorders caused by mutation in FGF receptor 3 (FGFR3) leading to Acondroplasia, the most common form of human genetic dwarfism. In October 2006, I joined the Institute of Biochemistry and Nutrition at the Faculty of Agriculture of the Hebrew University.
During these years, we (see the list of the lab, young talented students) uncovered new roles for FGF in chondrocytes and in mutant mouse models. We described the interactions between the cytokine receptor family, and FGFR signaling pathway. I also initiated a new topic of research into the role of matrix metalloproteases (MMPs) in skeleton development. The MMPs are a family of enzymes that are involved in proteolytic cleavage of matrix components. These enzymes can be modulated by diet (Ca, Zn, Cu) and are subject to complex transcriptional regulation, indicating a genetic-nutritional interrelation during bone development. We showed, for the first time, the perivascular localization of cMMP9 and proved its role in vascularization of the avian GP. This project was conducted by several students in my lab and generated several publications, grants, and synergistic international collaborations. In collaboration with Dr. Dalit Sela-Donenfeld, we questioned whether by degrading the extracellular matrix, the MMPs mediate delamination and migration of neural crest cells from the neural tube.
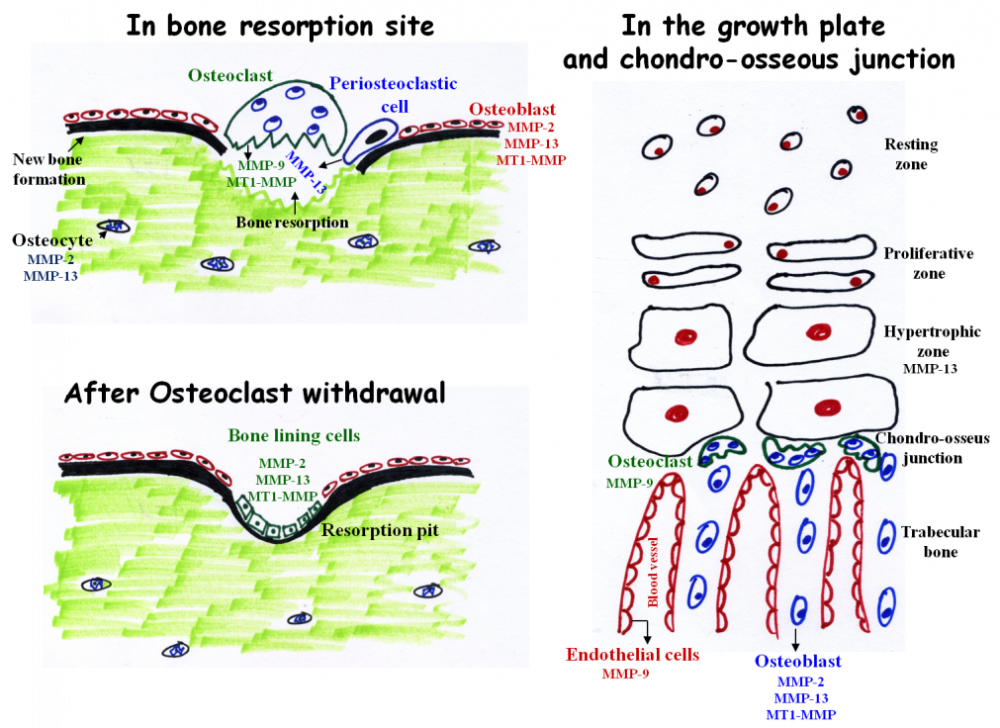
The main MMPs involved in bone resorption and endochondral ossification. During bone resorption MMP-9 and MT1-MMP are secreted by osteoclasts. However, these enzymes do not participate in matrix resorption per se but rather in osteoclasts migration and invasion. In contrast, MMP-13 is secreted from peri-osteoclastic osteocytes, translocate from the osteocytes into the resorption site where it participate in resorption. After osteoclast withdrawal, bone lining cells enter the lacuna and secrete MMPs (probably MMP-2, 13, and MT1-MMP) which remove demineralized collagen that was not digested by the osteoclasts. This “cleaning” is prerequisite for the subsequent deposition of a first layer of collagenous proteins in the resorption pit. MMP-9 and MMP-13 are the most important MMPs in endochondral ossification. MMP-13 is secreted from hypertrophic chondrocytes and MMP-9 from osteoclasts and endothelial cells

A continuing focus of interest of my lab is the effect of mechanical loading on the process of bone development. This is a multifaceted project, combining an in-vivo approach, in which we apply external loads on young fast-growing bones, with cell biology methods to understand the transduction of mechano-signaling in chondrocytes. Using various validation techniques, we identified load-dependent modifications in the major signaling pathways in the growth plate. These modifications reduce the number of chondrocytes in the GP, and therefore the latter's width and the final length of the bone. The possible involvement of the cilia in transducing mechano-signaling was further studied in tissue cultured chondrocytes by confocal microscopy and gene knock-down methods combined with loading protocols of cells in silicon chambers. This project involve collaboration with Prof. Nicolai Miosge dealing with the role of mechanical forces in tissue engineering of cartilage and mesenchymal cells. Within this research we also studied the effects of loading on bone phenotype in terms of architecture and mechanical properties. This research is carried out in collaboration with Prof. Ron Shahar, and has enabled us to purchase a micro-CT that significantly expands our scientific capabilities, combining our diverse expertise and joint interest in bone biology.
The broad aim of my research is to understand the role of nutritional and metabolic factors in bone development. The connection between under-nutrition and growth retardation is well documented. A trend of increasing height is recognized in countries where proper nutrition, exercise and fitness are more equally distributed. Children from developing countries with Protein-energy malnutrition (PEM) had significantly lower body weight and height than healthy subjects. Furthermore, animal studies clearly demonstrated the deleterious effects of PEM on linear growth. But what is the reciprocal? What if you eat too much? Are you going to over grow? Yes, it is recognized that overweight children are taller than age-matched, normal-weight children. This tall stature is often associated with an accelerated skeletal maturation and an earlier timing of puberty, but not with the final adult height. Thus it is clear that factors such as vitamin D and calcium are just the tip of the iceberg, and the role of nutritional factors in bone development has yet to be elucidated.
In the last few years, the connection between an organism's metabolic status and mature skeletal homeostasis has revolutionized the field of bone research, revealing the effect of leptin on bone density, and suggesting an endocrine loop in which a bone-specific protein (osteocalcin) affects insulin secretion and sensitivity. Despite these advances, other predictable link between organism's metabolic status and skeletal development has not been addressed. My research vision and activities aim to fill this gap. In this line, the projects we are conducting in the lab are dealing with:
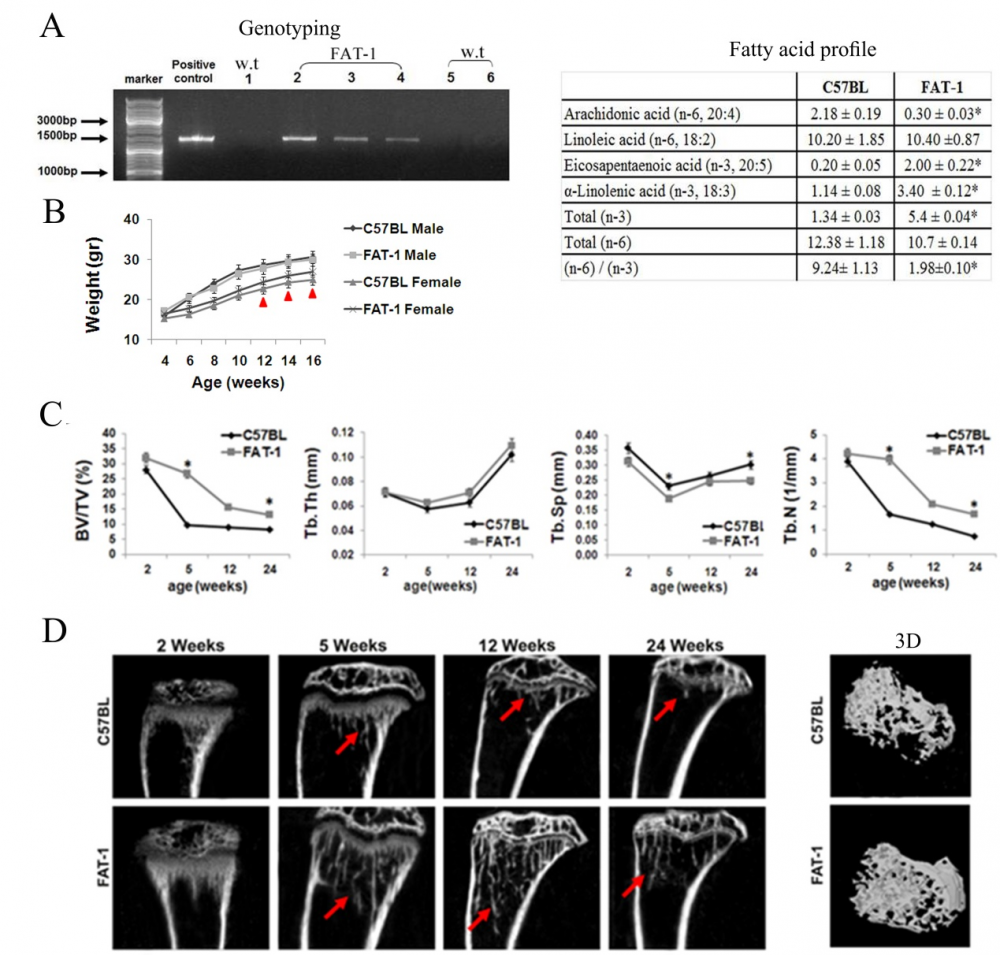
The effect of different fatty acids on bone development. We have shown the role of W-3 in the processes of bone modeling and remodeling, and its positive effects in the process of young bone development. For this project, in collaboration with Prof. Betty Schwartz, we are using the fat-1 transgenic mice, which are genetically modified to endogenously synthesize W-3 PUFA. We further studying the differential effects of high fat diets based on SFA, MUFA or PUFA on bone elongation and quality, and on the metabolic consequences of these diets. Our results suggest that these fatty acids are not only energy sources but may serve as signaling molecules in bone cells.
The effects of cytokines secreted from the adipose tissue on bone development and properties. In this project we are studying the effect of fat-derived cytokines on skeletal phenotype in-vivo by local or peripheral injections and by using mice models and in-vitro by chondrocytic or osteoblastic cell lines. By this combined approach we will get a new insight to the cross-talk between the adipose the cartilaginous and bone tissues during developmental stages. So far we have studied the role of leptin and adiponectin on embryonic bone development, and initiated an in-vivo model of obesity and metabolic syndrome during the early postnatal period by blocking the signaling of leptin.

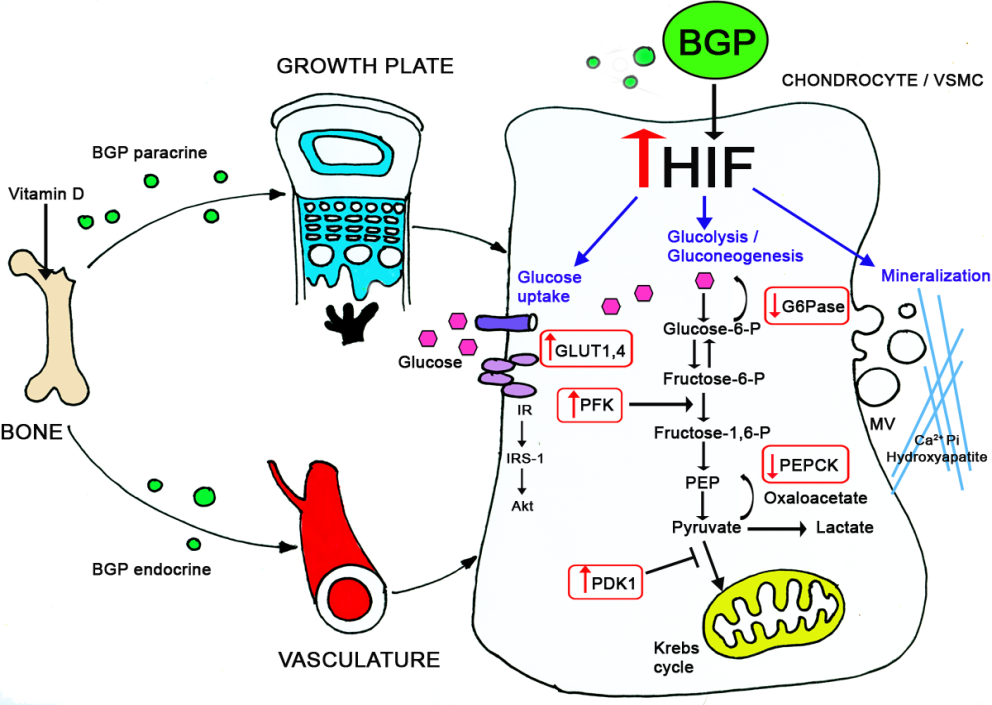
In a fascinating project we compared the plasticity of different mesenchymal cells in terms of differentiation and mineralization. Taking advantage of our expertise in chondrogenesis we were able to demonstrate that vascular smooth muscle cells undergo similar differentiation routs which results in the pathological condition of vascular calcification. We conducted a series of experiments in young rats (during their growth period) inducing arteries calcification by calcitriol injections. Using this model, and tissue culture approaches, we demonstrated the involvement of glucose metabolism in the physiological (in cartilage) and pathological (in vasculature) calcification. We further elucidated the underlying mechanisms regulating these events (through BGP and HIF-1α) by knocking down these genes in-vitro and in-vivo, using the Si-RNA approach.
In our most recent project we explore the effect of the western diet on bone development. In recent years obesity has become a public health crisis in the western society has tripled. As children's body weights have increased, so has their consumption of fast foods and soft drinks. The proportion of foods that children consume from restaurants and fast food outlets increased by nearly 300% between 1977 and 1996. Children’s soft drink consumption has also increased during those years. Junk foods, fast foods and snack foods are high in fat and refined sugars and don’t provide appropriate levels of fiber, protein, vitamins and minerals for the growth requirement of growing children. Thus, beside obesity, diet based on junk food may cause malnutrition. We examine the influence of childhood obesity and malnutrition, caused by unhealthy and unbalanced diet on the development and quality of the post-natal bone while the main focus is on the growth plates, as well as on the metabolic state of the body. We use young rats as our experimental model, and discovered that rats fed junk food diet, had impairments in bone growth and structure, which could result from the deficiency of nutrients in the junk food diet.
The metabolic regulation of skeletal development is of great importance from both basic and clinical points of view. From a practical standpoint, these concerns are related to the dramatic rise in childhood obesity, which is now reaching epidemic proportions. From a more basic standpoint, the fact that adipocytes, chondrocytes and osteoblasts all originate from the same progenitors (mesenchymal stem cells) and are able to trans-differentiate into one another, raises enthralling scientific possibilities. Just imagine that by manipulating these progenitor cells, ideally via nutritional or mechanical strategies, we might shift their programmed differentiation from fat-forming to bone-forming cells: if this vision would become reality, we could help solve two major health concerns in the world today, obesity and osteoporosis.
Curriculum Vitae
| 1987-1990 | B.Sc. degree in Human Nutrition (Cum-Laude), at the Hebrew University of Jerusalem, Faculty of Agriculture, Rehovot, Israel. |
| 1991-1997 | Direct studies towards Ph.D. at Volcani Center and the Faculty of Agriculture, the Hebrew University, Rehovot, Israel, Departments of Biochemistry and Animal Science. |
| 1997-2000 | Postdoctoral fellow at the Weizmann Institute of Science. Rehovot, Israel, Department of Molecular Cell Biology. |
| 2000–2006 | Research Scientist at the Agricultural Research Organization (ARO), The Volcani Center, Institute of Animal Science |
| 2005-2006 | Sabbatical at the ANZAC Research Institute; Sydney, Australia Bone Biology group. |
| 2006-2013 | Senior lecturer, Institute of Biochemistry, Food Science and Nutrition, The Robert H. Smith Faculty of Agriculture, Food and Environment. |
| 2013- | Associate Professor, Institute of Biochemistry, Food Science and Nutrition, Field: The Robert H. Smith Faculty of Agriculture, Food and Environment. Nutritional aspects of Physiology and Pathology of Bone Development. |
Teaching
Human Anatomy and Physiology, undergraduate students.
Endocrinology, undergraduate students.
Signal Transduction Pathways, graduate students.
Lab members
|
|
Efrat Monsonego-Ornan : PhD, PI |
|
In 2006, I joined the Institute of Biochemistry and Nutrition. This was a significant step for me, allowing me to fulfill my goal of conducting a research on the interrelation of genetic and environmental factors (such as nutrition) in bone development. I have always felt that the fast-developing and emerging field of nutrition is a fascinating discipline in terms of both research and public health. Furthermore, in joining the School of Nutrition I have in essence come full circle, since my first degree and in fact, first interest was nutrition. I therefore consider my current appointment both a mission and a privilege: I am able to study nutrition at the highest scientific level, in my research. And in teaching future nutritionists, I have the opportunity to highlight and enhance the scientific side of this crucial occupation. |
|
|
|
Svetlana Penn- Lab Technician |
|
|
Rotem Kalev: PhD Student |
|
|
Jerome Janssen: PostDoc |
|
|
Astar Shitrit: MSc Student |
|
|
Hodaya Godreich: MSc Student
|
|
|
Dana Meilich: MSc Student |
|
Seman Daeem: MSc Student |
|
|
|
Gal Becker: Amirim student |
|
|
Roni Sides: MSc Student |
|
|
Reut Rozner: MSc Student |
|
|
Janna Zaretsky: RD, PhD Student |
|
|
Shelley Griess: RD, PhD Student |
|
My name is Shelley, I'm a registered dietitian, who loves to cook. My great passion for the field of nutrition, and my unstainable curiosity drive me in my research. |
|
|
|
Ido Kedem: MSc Student |
|
|
Tamara Travinsky: RD. MSc Student |
|
Since I was in high school I was very interested in food and nutrition. My curiosity brought me to our laboratory and at the moment I am researching the effects of early nutrition on skeletal development. My curiosity is not even close to being satisfied and I truly hope it never will! |
|
|
|
Lior Shehav: Amirim Student |
|
|
Tali Sinai: RD. PhD. PostDoc |
















| Romi Nir | |
| Orit Ofir | |
| Gal Rub |
Former Students:
2001-2003 Ailing Tong
2001-2009 Dr. Adi Reich
2002-2004 Shimrit Weizmann
2002-2004 Tal Ben Zvi
2003-2005 Mira Negev
2003-2013 Dr. Stav Simsa
2003-2007 Ety Yael
2004-2007 Harel Dan
2004-2007 Marina Dolgin
2006-2008 Jenia Kosonovsky
2007-2011 Dr. Anna Idelevitch.
2007 Dr. Michal Ben-Ami
2008-2009: Dr. Tengana Dellesa
2008-2010: Lee Roth
2008-2011: Yoach Rais
2008-2011: Nur Abu-Ahmad
2008-2012: Olga Salinikov
2008-2013: Dr. Gili Solomon
2008-2012: Netta Ohayun
2009 -2011: Vietriki Fraggiranki
2009-2011: Tal Kfir
2010 Dr. Adi Ron.
2011-2012: Michal Har-Tal
2011-2012: Sergei Anpilov
2011-2014 Yeela Helman
2011-2014 Dafna Busani
2011- 2014 Lihi Morglander
2012- 2013 Rabindra Dithal
2013- 2014 Adi Carmi
2013- 2014 Maya Moshe
List of Publications
- Zaretsky J, Griess-Fishheimer S, Carmi A, Travinsky Shmul T, Ofer L, Sinai T, Penn S, Shahar R, Monsonego-Ornan E. (2021) Ultra-processed food targets bone quality via endochondral ossification. Bone research 9 (1), 1-13
- Kalev-Altman R, Hanael E, Zelinger E, Blum M, Monsonego-Ornan E, Sela-Donenfeld D . (2020) Conserved role of matrix metalloproteases 2 and 9 in promoting the migration of neural crest cells in avian and mammalian embryos. FASEB J. 34(4):5240-5261.
- Antunes BP, Vainieri ML, Alini M, Monsonego-Ornan E, Grad S, Yayon A. (2020) Enhanced chondrogenic phenotype of primary bovine articular chondrocytes in Fibrin-Hyaluronan hydrogel by multi-axial mechanical loading and FGF18. Acta Biomater.105:170-179.
- Rozner R, Vernikov J, Griess-Fishheimer S, Travinsky T, Penn S, Schwartz B, Mesilati-Stahy R, Argov-Argaman N, Shahar R, Monsonego-Ornan E. (2020) The Role of Omega-3 Polyunsaturated Fatty Acids from Different Sources in Bone Development. Nutrients. 12(11):3494.
- Ofir O, Buch A, Rouach V, Goldsmith R, Stern N, Monsonego-Ornan E. (2020) Association between abdominal obesity and fragility fractures among elderly Israeli women. Aging Clin Exp Res. 32(8):1459-1467.
- Sinai T, Goldberg MR, Nachshon L, Amitzur-Levy R, Yichie T, Katz Y, Monsonego-Ornan E, Elizur A.J. (2019) Reduced Final Height and Inadequate Nutritional Intake in Cow's Milk-Allergic Young Adults. Allergy Clin Immunol Pract. 7(2):509-515.
- Ofer L, Dean MN, Zaslansky P, Kult S, Shwartz Y, Zaretsky J, Griess-Fishheimer S, Monsonego-Ornan E, Zelzer E, Shahar R. (2019) A novel nonosteocytic regulatory mechanism of bone modeling. PLoS Biol. ;17(2):e3000140.
- Roth L, Kalev-Altman R, Monsonego-Ornan E, Sela-Donenfeld D. (2017) A new role of the membrane-type matrix metalloproteinase 16 (MMP16/MT3-MMP) in neural crest cell migration. Int J Dev Biol. 61(3-4-5):245-256.
- Kalish‐Achrai N, Monsonego‐Ornan E, Shahar R. (2017) Structure, composition, mechanics and growth of spines of the dorsal fin of blue tilapia Oreochromis aureus and common carp Cyprinus carpio. Journal of fish biology 90 (5), 2073-2096.
- Rub G, Marderfeld L, Poraz I, Hartman C, Amsel S, Rosenbaum I, Pergamentzev-Karpol S, Monsonego-Ornan E, Shamir RJ. (2016) Validation of a Nutritional Screening Tool for Ambulatory Use in Pediatrics. Pediatr Gastroenterol Nutr. 62(5):771-5.
- Dean MN, Ekstrom L, Monsonego-Ornan E, Ballantyne J, Witten PE, Riley C, Habraken W, Omelon S. (2015) Mineral homeostasis and regulation of mineralization processes in the skeletons of sharks, rays and relatives (Elasmobranchii). Semin Cell Dev Biol. 46:51-67.
- Solomon G, Atkins A, Shahar R, Gertler A, Monsonego-Ornan E. (2014) Effect of peripherally administered leptin antagonist on whole body metabolism and bone microarchitecture and biomechanical properties in the mouse. Am J Physiol Endocrinol Metab. 306:E14-27.
- Koren N, Simsa-Maziel S, Ron Shahar R, Schwartz B, and Monsonego-Ornan E. (2014). Exposure to omega-3 fatty acids at early age accelerate bone growth and improve bone quality. J Nutri Biochem. 25:623-33.
- Schminke, B. Muhammad, H. Bode, C. Boguslawa, S. Gerstdorf, N. Buergers, R. Monsonego Ornan, E Rosen, V. and Miosge, N. (2014). A discoidin domain receptor 1 knock-out mouse as a novel model for osteoarthritis of the temporomandibular joint. Cell Mol Life Sci.7:1081-96.
- Rais Y, Reich A, Simsa-Maziel S, Moshe M, Idelevich A, Kfir T, Miosge N, Monsonego-Ornan E. (2014) The growth plate's response to load is partially mediated by mechano-sensing via the chondrocytic primary cilium. Cell Mol Life Sci.
- Pando R, Masarwi M, Shtaif B, Idelevich A, Monsonego-Ornan E, Shahar R, Phillip M, Gat-Yablonski G. (2014) Bone quality is affected by food restriction and by nutrition-induced catch up growth. J Endocrinol. JOE-14-0486.
- Simsa-Maziel S, Reich A, Koren Y, Shahar R. and Monsonego Ornan E. (2013) L1-RI participates in normal growth plate development and bone modeling. Am J Physiol-Endoc M. 305(1):E15-21.
- Simsa-Maziel S, Sela-Donenfeld D. and Monsonego Ornan E. (2013) Matrix metalloproteinases in bone health and disease. Proteases in Cellular Dysfunctions, Springer.
- Bar A, Shoval I, Monsonego Ornan E and Sela-Donenfeld. (2013) The Role of Proteases in Embryonic Neural Crest Cells. Proteases in Cellular Dysfunctions, Springer.
- Simsa-Maziel, S. and Monsonego Ornan, E. (2012) Interleukin-1β promotes proliferation and inhibits differentiation of chondrocytes through a mechanism involving down-regulation of FGFR-3 and p21. Endocrinology. 153:2296-310.
- Monsonego Ornan, E. Kosonovsky, J. Bar, A. Roth, L. Renki-Fragis, V. Simsa-Maziel, S. Kohl A. and Sela-Donenfeld, D. (2012) Matrix Metalloproteinase 9/Gelatinase B is Required for Neural Crest Cell Migration. Dev Biol. 364:162-77.
- Cohen L, Dean M, Shipov A, Atkins A, Monsonego-Ornan E. and Shahar R. (2012) Comparison of structural, architectural and mechanical aspects of cellular and acellular bone in two teleost fish. J Exp Biol. 215:1983-93.
- Amosi N, Zarhitsky S, Salnikov O, Monsonego-Ornan E, Shahar R. and Rapaport, H (2012) Acidic peptide hydrogels loaded with β-TCP particles act synergistically to induce bone regeneration. Acta Biomaterialia.8:2466-75.
- Simsa-Maziel, S, Reich, A, Sela-Donenfeld, D. and Monsonego Ornan E. (2012). The role of Matrix Gla Protein in ossification and recovery of the avian growth plate. Frontiers in Bone Research; in Frontiers in Endocrinology.3:79. Dan, H,
- Muhammad, H. Rais, Y. Miosge, N. and Monsonego Ornan, E. (2012) The primary cilium as a dual sensor of mechanochemical signals in chondrocytes. Cellular and Molecular Life Sciences. 69(13):2101-7.
- Weisinger, K., Kohl, A., Kayam, G., Monsonego-Ornan, E and Sela-Donenfeld, D. (2011) Expression of hindbrain boundary markers is regulated by FGF3. Biology Open. 1:67-74.
- Bar-El Dadon S., Shahar, R., Katalan V., Monsonego-Ornan, E. And Reifen R. (2011) Leptin administration affects growth and skeletal development in a rat intrauterine growth restriction model: Preliminary study. Nutrition. 27:973-7.
- Idelevich, A, Kerschnitzki, M, Shahar, R. and Monsonego-Ornan, E. (2011) 1,25(OH)2D3 alters growth plate maturation and bone architecture in young rats with normal renal function. PlosOne. 6, e20772.
- Sharir A, Israeli D, Milgram J., Monsonego-Ornan E., Currey JD. and Shahar R. (2011) The canine baculum: the structure and mechanical properties of an unusual bone. J Structural Biol. 175: 451.
- Idelevich, A., Rais, Y. and Monsonego-Ornan, E. (2011) Bone Gla Protein increases HIF-1α-dependent glucose metabolism and induces cartilage and vascular calcification. Arterioscler. Thromb. Vasc. Biol. 31:e55-71.
- Reich, A., Simsa-Maziel, S., Ashkenazi, Z. and Monsonego Ornan E (2010) Involvement of matrix metalloproteinases in the growth plate response to physiological mechanical load. J Appl Physiol. 108(1):172-80.
- Delessa Challa, T, Rais, Y and Monsonego Ornan, E (2010) Effect of Adiponectin on ATDC5 Proliferation, Differentiation and Signaling Pathways. Mol Cell Endocrinol. 323:282-91.
- Dan, H., Simsa-Maziel, S., Hasdai, A., Sela-Donenfeld, D. and Monsonego Ornan E. (2009) Expression of matrix metalloproteinases during impairment and recovery of the avian growth plate. J Anim Sci.87:3544-55.
- Shipov, A, Sharir, A, Zelzer, E, Milgram, J, Monsonego Ornan, E. and Shahar, R. (2009) The influence of severe and prolonged exercise restriction on the mechanical and structural properties of bone: an avian model. Vet J. 18, 153-160.
- Reich A., Sharir A., N., Zelzer E., O., Hacker L., Monsonego Ornan E. and Shahar R. (2008) The effect of weight loading and subsequent unloading on the post natal skeleton. Bone. 43:766-74.
- Negev-Hasky M., Simsa S., Tong A., Genina O. and Monsonego Ornan E. (2008)Involvement of the matrix metalloproteinases in vascularization and ossification of normal and impaired avian growth plate. J Anim Sci.86:1306-15.
- Monsonego Ornan E, and Ben-Ami M. (2008) The role of leptin in bone development and growth. Landes Bioscience. Leptin and Leptin Antagonists. 83-90.
- Krauss, S., Monsonego Ornan, E., Zelzer, E., Fratzl, P. and Shahar, R. (2008) Mechanical function of a complex three-dimensional suture joining the bony elements in the shell of the red-eared slider turtle. Advanced Material. 21, 407–412.
- Simsa, S. and Monsonego Ornan, E. (2007) Endochondral Ossification Process of the Turkey (Meleagris gallopavo) During Embryonic and Juvenile Development. Poult Sci. 86:565.
- Simsa S, Hasdai A, Dan H and Monsonego Ornan E.(2007) Differential regulation of MMPs and matrix assembly in chicken and turkey growth-plate chondrocytes. Am J Physiol Regul Integr Comp Physiol; 29:2216-24.
- Simsa S, Genina O, Monsonego Ornan E. (2007) Matrix Metalloproteinase expression and localization in turkey (Meleagris gallopavo) during the endochondral ossification process. J Anim Sci.;85:1393-401.
- Simsa, S., Hasdai, A., Dan, H. and Monsonego Ornan, E. (2007) Induction of Tibial Dyschondroplasia in Turkeys by Tetramethylthiuram Disulphide (Thiram). Poult Sci.86:1766-71.
- Leach Jr. RM. and Monsonego Ornan E. (2007). Tibial dyschondroplasia – tools, new insights and future prospects World Poul. Sci. 86:2053-58.
- Ben-Zvi T, Yayon A, Gertler A and Monsonego Ornan E (2006). Suppressors of cytokine signaling (SOCS) 1 and 3 interact with and modulate fibroblast growth factor receptor signaling. J Cell Sci. 119:380-7.
- Reich A, Jaffe N, Genina O, Lavelin I, Nussinovitch A and Monsonego Ornan E (2005) Weight loading on young chicks inhibits bone elongation and promotes growth plate ossification and vascularization. J app Physiol. 98:2381-9.
- Weitzman S, Tong A, Reich A, Genina O, Yayon A and Monsonego Ornan E (2005). FGF upregulates osteopontin in epiphyseal growth plate chondrocytes: Implications for endochondral ossification. Matrix Biol 24:520-9.
- Tong A, Reich A, Genin O, Pines M and Monsonego Ornan E (2003) The expression pattern of chicken 75-kDa gelatinase B-like enzyme in chondrocytes: evidence for its role in vascularization of the growth plate. J Bone Miner Res 18:1443-52.
- Rozenblatt-Rosen O., Monsonego Ornan E., Sadot E., Madar-Shapiro L., Sheinin Y., Ginsberg D. and Yayon A. (2002) Induction of chondrocyte growth arrest by FGF: transcriptional and cytoskeletal alterations. J Cell Science 115; 553-62.
- Monsonego Ornan E, Adar R, Rom E and Yayon A (2002) Ligand-mediated ubiquitylation of wild type and mutant FGF receptors: dependence on tyrosine kinase activity and role in downregulation. Febs Letters. 528:83-9.
- Eswarakumar V.P., Monsonego Ornan E., Pines M., Antonopoulou I., Gillian M., Morriss-Kay and Lonai P. (2002) The IIIc alternative of Fgfr2 is a positive regulator of endochondral ossification. Development. 129: 3783-93.
- Adar. R., Monsonego Ornan E. and Yayon A. (2002) Effects of cysteine mutations in fibroblast growth factor receptor 3 on receptor dimerization and activation. J.Bone.Miner.Res 17: 860-8.
- Monsonego Ornan E, Adar R, Feferman T, Segev O and Yayon A (2000) The Achondroplasia mutation G380R in FGFR3 uncouples receptor activation from internalization. Mol Cell Biol 2; 516-22.
- Chen L., Adar R., Yang X., Monsonego Ornan E., Li C., Yayon A. and Deng C. (1999) An Achondroplasia type mutation (G369C) in murine FGFR3 retards long bone growth by inhibiting both chondrogenesis and osteogenesis. J. Clinical Invest. 104: 1517-25.
- Monsonego, E., Baumbach, W, Lavelin, I., Gertler, A, Hurwitz, S., and Pines, M. (1997). Generation of growth hormone binding protein by avian growth plate chondrocytes is dependent on cell differentiation. Mol Cell Endocrinol 135, 1-10.
- Pines, M., Monsonego, E., Knopov.V., Shalom-Barak, T, Halevy, O., and Hurwitz, S. (1995). The involvement of the epiphyseal growth-plate in longitudinal bone growth and in tibial dyschondroplasia. Archiv Fur Geflugelkunde. 28-30.
- Monsonego, E., Halevy, O., Gertler, A., Hurwitz, S., and Pines, M. (1995). Growth hormone inhibits differentiation of avian epiphyseal growth- plate chondrocytes Mol Cell Endocrinol 114, 35-42.
- Halevy, O., Monsonego, E., Marcelle, C., Hodik, V., Mett, A., and Pines, M. (1994). A new avian fibroblast growth factor receptor in myogenic and chondrogenic cell differentiation. Exp Cell Res 212, 278-84.
- Monsonego, E., Halevy, O., Gertler, A., Volokita, M., Schickler, M., Hurwitz, S., and Pines, M. (1993). Growth hormone receptors in avian epiphyseal growth-plate chondrocytes. Gen Comp Endocrinol 92, 179-88.
- Pines, M., Monsonego, E., Shalom-Barak, T., Halevy, O., and Hurwitz, S. (1993). Effect of hormones and growth factors on cell proliferation and synthesis of extracellular protein by epiphyseal growth-plate chondrocytes. Avian Endocrinology Ed. P.J. Sharp 249-260.
From the Media
ערוץ 13, נגה ניר-נאמן, ״דופק את הגוף": כך משפיע הג'אנק פוד על ההתפתחות של ילדינו
ערוץ 13, נגה ניר-נאמן, אל תאמינו לאריזות
כינוס של מכון ואן ליר ״זה הזמן לפוסטקפיטליזם״.
מושב מיוחד בנושא תזונה בעידן הפוסטקפיטליסטי: הדרך ליצירת משק תזונתי בריא, דמוקרטי ומקיים
החקולטה- הפודקאסט של הפקולטה לחקלאות
פרק 4- ב': המשך השיחה עם פרופ אפרת מונסונגו אורנן
פרק 4- א': מזון אולטרא מעובד וההשפעה שלו על הגוף שלנו
עיתונות כתובה
"יש לכן פצצה ביד": הכירו את האוכל שמפורר לילדינו את העצמות
دراسة تكشف خطورة الوجبات السريعة على عظام الأطفال
La malbouffe met la santé osseuse des enfants en danger
Caution for Kids: Junk Foods are Bad to the Bones
Researchers reveal the effect of ultra processed foods on skeleton development
Bad to bone: Hebrew University reveals impact of junk food on kids’ skeletal development
Bad to the bone: Junk food damages childhood bone development, study finds































